While this efficiency is impressive, it also is rarely achieved. The difficulty is in the protein that carries out the first step of photosynthesis. Molecular oxygen, O 2, competes with CO 2 for the active site of ribulose bisphosphate carboxylase, leading to an oxidation and loss of the ribulose bisphosphate acceptor. This competition is apparently intrinsic to the enzyme, because attempts to increase the discrimination for CO 2 by genetic engineering have resulted in a less‐active enzyme, which fixes CO 2 very poorly.
Oxidation of ribulose‐1,5‐bisphosphate by Rubisco produces a 3‐carbon compound, 3‐phosphoglycerate, and a 2‐carbon compound, phosphoglycolate. Because carbon is oxidized, the process is termed photorespiration. ![]()
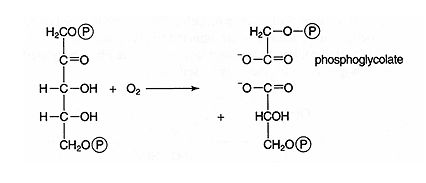
Photorespiration reduces the efficiency of photosynthesis for a couple of reasons. First, oxygen is added to carbon. In other words, the carbon is oxidized, which is the reverse of photosynthesis—the reduction of carbon to carbohydrate. Secondly, it is now necessary to resynthesize the ribulose bisphosphate and to reduce the phosphoglycolate.
The 3‐phosphoglycerate from photorespiration can reenter the Calvin‐Benson pathway, but the phosphoglycolate must be recycled to make a useful compound. This recycling takes place in a specialized organelle termed the peroxisome. Peroxisomes lie between chloroplasts and mitochondria in the plant cell and serve to pass the 2‐carbon products of oxygenation on for further metabolism. In the chloroplast, the phosphoglycolate is dephosphorylated. Glycolate is transported to the peroxisome where molecular oxygen further oxidizes it to glyoxylate. The product is hydrogen peroxide, H 2O 2, (the term peroxisome comes from this product) which is rapidly broken down by catalase to water and oxygen.
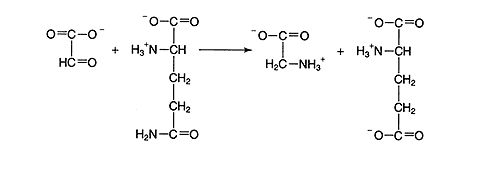
The glyoxylate is amidated to the amino acid glycine in the peroxisome.
Glycine is then transported to the mitochondrial matrix where the conversion of two glycines to one serine occurs with the loss of CO 2 and NH 3 from the pool of fixed molecules. The serine is transported into the peroxisome, where it is deaminated to glycerate. The glycerate is transported back to the chloroplast, where it is phosphorylated to 3‐phosphoglycerate for the Calvin‐Benson cycle. ![]()
This set of reactions is very detrimental to the efficiency of photosynthesis. Oxygen is added to carbon, CO 2 is lost, energy is consumed, and ribulose bisphosphate is destroyed. For a plant to be able to increase the discrimination of Rubisco for CO 2 would obviously be advantageous, but that hasn’t happened, either naturally or through the efforts of scientists. An increased concentration of CO 2 in the atmosphere may lead to increased photosynthesis and decreased photorespiration, but high CO 2 concentrations would also contribute to global warming (and the increased photosynthetic carbon fixation would not likely reduce the amount of CO 2 in any event).

Figure 1
C4 plants
Photorespiration could be overcome if Rubisco could receive CO 2 more efficiently than by diffusion through the leaves. Various C4 plants, including grasses such as maize (corn) and bamboo, have evolved a bypass system for the delivery of CO 2 to Rubisco.
In these C4 plants, the enzyme phosphoenolpyruvate carboxylase first converts CO 2 to oxaloacetate. ![]()
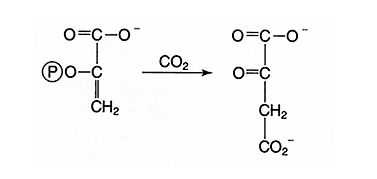
PEP carboxylase has a lower K m for CO 2 than does Rubisco. Further, O 2 is a very poor substrate for this enzyme. This means that, at relatively low concentrations of CO 2, the delivery of carbon into photosynthesis products is more efficient than in C3 plants and oxygenation doesn’t occur.
PEP carboxylase is concentrated in special mesophyll cells in the outer part of the leaf. This means that the cells most exposed to the atmosphere are the most efficient at converting CO 2 into organic products. Photosynthesis involving Rubisco is more prominent in the bundle sheath cells located in the inner part of the leaf around the veins that carry compounds between different parts of the plant.
After PEP carboxylase makes the oxaloacetate, it is transported to the bundle sheath cells. First, NADPH reduces it to malate, and it is then transported to the bundle sheath cells. In the bundle sheath cells, malic enzyme cleaves the malate to pyruvate and CO 2 for Rubisco. This generates NADPH as well, so the C4 cycle consumes no reducing equivalents. Pyruvate is transported from the bundle sheath back to the mesophyll cells where it is rephosphorylated to phosphoenolpyruvate, expending the equivalent of two ATP “high‐energy phosphates.”
Overall, the C4 cycle consumes two ATP equivalents to deliver a CO 2 to Rubisco. During active photosynthesis, this is not a problem—plenty of ATP exists from the action of Photosystems I and II. Why, then, don’t C4 plants take over the world? Probably because the increased energy demands make these plants less efficient under conditions where sunlight is limited. Consistent with this idea, C4 plants are mostly confined to tropical climates, while the C3 plants predominate in more temperate regions.